|
The Evolutionary Theory of
Sex:
Variation of Sexes and Reaction Norm
 A
man marries a woman thinking she will never change A
man marries a woman thinking she will never change
and a woman marries a man thinking he will change.
And they are both wrong.
It was noted
already that males are first who die from all damaging
environmental factors. The male sex is a biologically weaker
sex. If we draw a mortality curve for each sex separately,
the male curve should contact with the front of the harmful
environmental factor. Hence, either curves are shifted, so
that the male curve is positioned between the front of the
harmful factor and a female curve, or male’s curve should
have a greater variation. The first decision is not
satisfactory, because rescuing a female from the given
harmful factor of environment (let’s say, cold) exposes them
to an alternative factor (in this case, heat). This leaves
us with the second decision—male’s curve should have
greater phenotypic variation than female’s curve
(Figure 1B). This is the corner stone of the new theory.
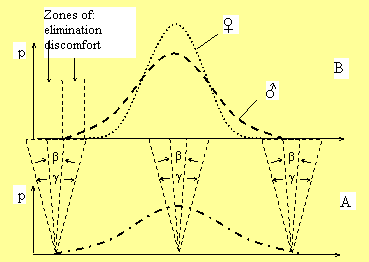
Figure
1
Distribution of male (- - -) and female (. . .)
genotypes (A) and phenotypes (B) in population.
β
and
γ—male
and female reaction norms accordingly.
The hypothesis
concerning wider phenotypic variation of males was
proposed in 1965. It permitted an easy explanation of many
earlier obscure phenomena, and first of all of the different
mortality of sexes (Geodakian, 1974). Higher mortality of
males is one of the puzzles of sexuality, a general
biological phenomenon, which no theory could explain
satisfactorily. In new theory it is interpreted as a
“payment” for new ecological information, as a useful form
for population to get new information from the environment.
For example males have higher susceptibility to all “new”
diseases of our century (infarction, arteriosclerosis,
cancer, schizophrenia and others).
What are the
causes and sources of this phenomenon? Why is the
phenotypic variation of males broader than that of
females?
Wider
phenotypic variation of males can be a consequence of
their wider genetic variation. It can also reflect
wider reaction norm of females which allows them to leave
zones of elimination and elimination discomfort. Wider
genetic
variation of males can be due to their higher
mutation rate. More additive inheritance of parental
characters by female offspring can decrease their variation
compare to males.
Wider Reaction Norm
of a Female
One more
phenomenon may bring about different phenotypic variation
of sexes—different reaction norm for males and females. The
norm of reaction, or modificational variability, individuals
of a female sex should be more than the appropriate reaction
norm of males. In other words, a male’s phenotype is more
“rigidly” related to genotype than a female’s. Therefore, in
Ontogeny, the environment has more influence upon
realization of a female genotype into a phenotype.
There is a certain zone of “ontogenetic discomfort” in
which individuals do not
perish, but experience inconveniences and difficulties,
suffer from lack of adaptation (freezing, starving and so
forth)
(Figure 1B). The wide reaction norm allows female individuals to
leave this zone and survive, while males stay in it and
perish. Female individuals have bigger “space of abilities”
due to their wide reaction norm compare to males. Therefore
the sex differentiation can be treated also as a
specialization on mutually additional qualities of
perfection (female sex) and innovation, progressiveness
(male one) in their relationship with the environment. The
adaptation of female individuals to existing environmental
conditions is more perfect, than males. On the contrary, the
presence of various deviations (imperfections) from the norm
(mode) of a population, give males more advantage in the
adaptation to changed conditions of environment in the
future.
The narrow
reaction norm of males provides them wide phenotypic
variation. Contrary, wide reaction norm of females narrows
their phenotypic variation. At a first glance it seems that
the wider reaction norm should result in wider phenotypic
variation. However it is not so. It is actually the
opposite: in a population where male phenotypic variation
consists of individuals with narrow reaction norm, it will
be wider, because it will be more close to their genotypic
variation. A female’s phenotypic distribution consists of
individuals with wide reaction norm. Therefore a female’s
phenotypic variation curve will be narrower, since in
conditions of stabilizing selection, the wide reaction norm
will allow extreme genotypes to “escape” zones of
“discomfort” and to come closer to phenotypic norm (mode) of
a population. High ontogenetic plasticity of a female sex
provides it’s high stability in Phylogeny. Hence, the
male sex is more flexible in the phylogenetic plan, and the
female sex is more flexible in the ontogenetic aspect. Such,
at first sight paradoxal, role distribution in Phylogeny
and Ontogeny actually is consecutive and logical. It
realizes uniform idea of specialization of sexes on
conservative and operative tasks of evolution. Slightly
exaggerating, it is possible to tell, that informational
relationship of a population with the environment is based
upon the elimination of males and the “education”
(ontogenetic shift) of females.
◄
Receiving the Ecological Information from the Environment
High Male Mortality and Gamete Type ►
High Male Mortality under extreme conditions leads to
decrease in tertiary sex ratio. Simultaneously more males
are required for selection. See
Sex
Ratio
►
|