|
Sex Chromosomes: What Are
They For?

Answers the question—What sex chromosomes are for?
Explains the routes of gene movement along the chromosomes
during the gene’s life-span.
A new concept uses the idea of
asynchronous evolution for the explanation of existence and
functional roles of sex chromosomes.
In 1965 V. Geodakyan theoretically
predicted that “In the chromosome set, the roles of sex
chromosomes and autosomes (A) is in effect, respectively, the
short-term and long-term memory; that is why sex
chromosomes, primarily the Y-chromosome, serve as a gateway
into heredity for variation”. Y-chromosome does it primarily
for mutations induced and directed by the ecological
differential rather than of spontaneous mutations.
The new genes first appear in the
Y-chromosome and then, after many generations, become
transmitted through the X-chromosome to autosomes.
Therefore, before appearing in
autosomes, each new gene is tested twice in sex
chromosomes, first in the Y- and then in the X-chromosome.
The recessive X genes manifest themselves almost exclusively
in males, and only males are under selection pressure. Such
is the evolutionary significance of sex differentiation and
the main advantage of sexual dimorphism.
The
Evolutionary Significance of Autosomes and Sex Chromosomes
Autosomes are long-term memory
of the genome (analogous to the female sex in a population),
and thereby serve for genome conservation. They store
the working structural genes common for both sexes. They
are the oldest chromosomes, found even in sexless
organisms. Autosomes contain fundamental species-specific
information, and implement the most ancient programs of
reproduction and recombination. Stochastic
transmission of autosomes ensures the maximum diversity of
genotypes. Owing to that, autosomes perfectly fulfill the
programs of the sexual process (fertilization), in which
remarkable achievements are made by hermaphrodites. In this
sense, autosomes can be regarded as “recombination”
chromosomes.
Sex chromosomes are short-term
memory structures, the experimental subsystem of the genome
(a male sex analogue), and provide for genome
modification. As a new character does not appear in the
female genotype without prior testing in the male one, a new
gene is not transferred to autosomes without testing in sex
chromosomes.
The main role of sex chromosomes is not
reproductive — determination of sex — as believed
earlier, but evolutionary, i.e., the creation of
dichronomorphism for economical evolution, even at the cost
of reproduction. Sex chromosomes change and
regulate the sex ratio rather than maintain it at
1:1 level.
Sex chromosomes are significantly
“younger” than autosomes. By triggering and implementing the
differentiation program, sex chromosomes form two
subsystems in a population (conservative and operative).
Because of this, the more monogamous and more polygamous
sexes have respectively broader and narrower reaction norms,
irrespective of the gametic type. Sex chromosomes contain
mainly evolving genes, both newly acquired and bound to be
lost. Their activity is directed against the
recombination program, as they restrict the male-male
and female-female combinations. Thus, the efficiency of the
sexual process is reduced twofold. In this sense, they are
more “antisex” rather than “sex” chromosomes. Therefore,
according to their role, it’s more logical to name them the
“evolutionary” chromosomes.
Algorithms
of information transmission and chromosomes
Several
algorithms can be identified in the informational behavior
of chromosomes.
Vertical mechanisms involve transmission of
chromosomes from generation to generation.
Horizontal mechanisms describe reception of
information from the environment; its transmission between
chromosomes; and elimination in the form of genetic
processes, such as mutagenesis, crossing-over,
translocations, episomal transfer, transfer by viruses,
plasmids, mobile elements, etc.
Behavior of the
chromosomes is determined primarily by three vertical
algorithms. (1) Stochastic, when chromosomes of the
homologous pair come to the son or daughter in a purely
random fashion. This algorithm is responsible for the
transmission of autosomes and thought to be typical for
X-chromosomes of the homogametic sex. (2) Ipsi-algorithm,
when the chromosome is transmitted from the parent only to
the offspring of the same sex. This is the transmission
algorithm of the Y-chromosome. (3) Contra-algorithm is
responsible for transmission from the parent to the
offspring of opposite sex. This is the transmission
algorithm of X-chromosome in the heterogamete sex.
The stochastic
algorithm deals with the genetic information that is common
for both sexes, and implies gene mixing during each
fertilization, which homogenizes the population, and hence
cannot provide a basis for genotypic sexual dimorphism. This
algorithm is the oldest. It existed even before sex
differentiation, and realizes only the reproduction and
recombination programs.
Nonstochastic
algorithms came into light after the appearance of different
sexes, and deal with the information which differs in males
and females, i.e., with genotypic sexual dimorphism; they
hence produce, maintain, and regulate the dimorphism. Ipsi
algorithms, pertaining to a single sex, transmit the genetic
information via males, and hence can produce genotypic
sexual dimorphism and increase or decrease it. Ipsi
algorithms initiate the differentiation program. Contra
algorithms, like the stochastic ones, involve information
transmission from one sex to another, hence homogenizing the
population, but contrastingly, they maintain the genotypic
sexual dimorphism at a stable level rather than reduce it to
zero. The combination of ipsi and contra algorithms thus
allows producing and maintaining a certain difference
between subsystems, and vary it in dependence on the
environmental condition. In this connection, contra
algorithms operate as stabilizers (negative feedback), while
ipsi algorithms operate as regulators (positive feedback).
The Y-chromosome is a connecting
link between the nucleus and cell environment (the
cytoplasm, mitochondria, etc.), a kind of genomic gateway
for information. This chromosome converts ecological
information into a genomic format, i.e., it produces new
genes (mutations), and can, therefore, be designated the
ecological chromosome. It contains genes “of tomorrow”;
triggers, accelerates, and controls the development of
asynchronism or sexual dichronomorphism (evolutionary and
morphological “distance” between sexes, including the
difference with respect to the reaction norm); and serves as
a primary “test range” and “quarantine” for new genes.
The contra-X chromosome (cX) is
a gene carrier linking the Y-chromosome with the female
genome (transport chromosome). Phylogenetically, it
functions as a stabilizer, relaxer, and liquidator of sexual
dichronomorphism. It also serves as the second test range
for new genes (hemizygous in males and heterozygous in
females) during ontogeny.
The ipsi-X chromosome (iX) is
involved in determining the reaction norm in females,
depending on the type of polygamy. It should contain a
greater proportion of modifier genes and genes encoding
quantitative traits. This chromosome transfers the new genes
to autosomes and eliminates their old genes. Its certain
region contains exclusively female genes “of yesterday,”
i.e., carries atavistic information. Thus, sex chromosomes
provide a means for both the storage and transmission of
information.
Evolutionary route of genes through chromosomes.
The phylogenetic pathway of information
transmission is as follows:
|
environment |
|
_______________
Chromosomes
_______________ |
|
↓ |
|
│ |
|
|
|
|
|
|
|
|
|
|
|
│ |
|
cytoplasm |
→ |
Y |
→ |
cXm |
→ |
cXf |
→ |
A |
→ |
iXf |
→ |
cXf |
→ |
cXm |
| |
Appearance of new genes
—> |
│ |
—>
Death of unnecessary genes |
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
m – male, f – female |
In the process of
partial conjugation of the X- and Y-chromosomes in animals,
just as in plants, only a part of genes is present in the
conjugating region (Fig). The "entry point" and the "exit
point" of the Y-chromosome should be at some distance from
each other. There should be some time necessary for the gene
to move along Y-chromosome in order to get to the region,
which conjugates with the X-chromosome.
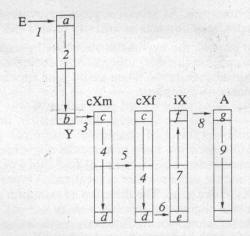
Fig.
The hypothetical schematic representation of the route of a
new gene through
sex chromosomes during the divergent
phase of trait evolution.
E—environment (cytoplasm). Chromosomal regions: a, c, e,
g —entries of chromosomes do not participate in
crossing-over; b, d, f —exits of chromosomes
participating in non-equal crossing-over and translocation
of genes to autosomes. Gene transitions: (1) from the
environment to Y chromosome (mutagenesis); (2) along
the Y chromosome from the nonconjugating (a) region
into the conjugating one (b); (3) nonequal
crossing over Y → cXm; (4) along the
length of cX chromosome in male and female genome; (5)
vertical cX algorithm (father → daughter); (6)
unequal crossing over cXm → iX; (7)
along the iX chromosome; (8) translocation
(plasmids, viruses) and iX chromosome → autosomes;
(9) along the length of autosomes.
The situation in
the X-chromosome appears to be similar to that in the
Y-chromosome. The entry point and the exit point are at some
distance from each other, and, therefore, moving along the
X-chromosome, the "young" gene in a hemizygous state is
subjected to a second testing period in male genome (Fig.).
Therefore, each new gene, before it moves into an autosome,
passes a double check in sex chromosomes: first, in the
Y-chromosome, then in the X-chromosome. But since recessive
X-genes are expressed only in males, it is the male sex that
is subjected to selection.
What
Genes Are Localized in Sex Chromosomes and in Autosomes ?
The classical chromosomal theory does
not give an answer to this question. However, the very name
of sex chromosomes (gonosomes) implicates that they carry
genes for characters associated with sex and reproduction,
and, consequently, autosomes carry genes for non-sexual,
somatic characters; i.e., genes are localized in chromosomes
according to the reproductive criterion. The new
concept gives a definite answer to the above question: sex
chromosomes must carry genes for evolving characters, and
autosomes, genes for constant (stable) ones; i.e., the
criterion is evolutionary.
Thus, according to the classic theory, somatic mutations
should be in autosomes, while genes coding for primary
sexual characters should be in sex chromosomes; now,
according to the new concept, all this should be the other
way around: somatic mutations should be located in sex
chromosomes, while genes coding for sexual characters should
be in autosomes (Table).
Table. Localization of genes according to the
classical vs. new theory.
|
Theory |
Traits |
|
stable |
evolving |
|
somatic |
sexual |
somatic |
sexual |
| Classic |
A |
SC |
A |
SC |
| New |
A |
A |
SC |
SC |
A —
autosomes, SC — sex chromosomes.
In the gene pool of a dioecious
population, genes can be classified into the following three
groups according to their evolutionary age and localization
in chromosomes of males and females:
(1) Exclusively male Y- and X-chromosome genes. These are
new, “young,” “tomorrow” genes determining future characters
that appeared in males but were not yet tested, transmitted
to autosomes, and shared.
(2) Shared (working, “today”) genes. These constitute the
main part of the genome, are localized in autosomes, and are
present simultaneously in both sexes.
(3) Female X-chromosome (old, having worked in autosomes,
“yesterday”) genes. These genes were already lost by males
but still preserved in females as atavistic prior to
elimination. These genes are probably localized in a special
region of the X chromosome (or maybe in autosomes) and are
transferred to the male genome only for elimination.
The condensation of the X chromosome in the female genome
The classic theory of sex chromosomes
interprets the condensation of the X chromosome in the
female genome (Barr's body), as the dose compensation
of X-chromosome genes. If this interpretation were true,
then the Barr bodies would normally always be present in
homogametic sex. However, in birds, as in mammals, the
female chromosome is condensed, although birds possess only
one X chromosome. How can this be explained on the basis of
dose compensation? Moreover, birds lack the conjugation of
sex chromosomes. For some unknown reason, DNA replication of
the single X chromosome in homogametic sex and the Y
chromosome occurs after the termination of autosome
replication.
According to the new concept the
condensation of the X chromosome in the female genome
irrespective of the gamety type, is interpreted as a
barrier to the spreading of new, untested
information in females.
From
mothers we receive old genetical information, from
fathers—latest evolution “news”
About two decades ago British scientists made an interesting
discovery. They showed that development of the placenta
(female organ) is controlled by father’s genes, while
development of an embryo—by mother’s [Surani, 1984, McGrath,
1984].
This
genetic process called genomic
imprinting is challenging the classical view of human
genetics that there are
two functioning copies, one from each parent, of every
gene. Imprinting means that one copy inherited from one
parent is active, for some genes the maternal copy and for
others the paternal, while the same gene from the other
parent is inactive or silent.
Genomic
imprinting confirms that there can be “male” and “female”
forms (or phases) of the same gene.
According to new view the main factor determining which form
is expressed is phylogenetic age of a character, system or
organ. Placenta is relatively new organ appeared in
placental mammals. Embryo is very old system. So, genes for
the placenta are still located in the male genome or were
moved into female genome but are still closed. Control of
embryonic development is very old information that we get
from mothers.
More about sex chromosomes
theory:
First publication:
Sex Chromosomes: What Are They For? (A New Concept).
Geodakyan V. A. Doklady Biological Sciences. Vol. 346,
1996. pp. 43-47. Translated from Doklady Akademii Nauk, Vol.
346, No. 4, 1996. pp. 565-569.
Popular version: N/A
Most complete scientific version:
The Role of Sex Chromosomes in Evolution: a new Concept.
Geodakian V. A. J. of Mathematical Sci. 1999, v. 93, № 4, p.
521–530.
Most recent publication:
Evolutionary Chromosomes And Evolutionary Sex Dimorphism.
Geodakyan V. A. Biology Bulletin, 2000, v. 27, № 2,
p. 99–113. Translated from Izvestija Akademii Nauk, Serija
Biologicheskaya, No. 2, pp. 133-148, 2000.
|