|
The Evolutionary Theory of
Sex: High Male Mortality and Gamete Type
 In a course of Ontogeny the sex
ratio for many species of plants, animals and humans goes
down. It is related to the raised death rate and
damageability of male’s systems in comparison with female
ones at almost all Ontogeny stages and at all levels of
organization. Whether we study various species (the humans,
animals or plants), different levels of the organization (an
individual, organ, tissue or a cell) or stability to
different harmful factors of environment (low and high
temperature, starvation, poisons, parasites, diseases,
etc.)—anywhere the same picture is observed: the raised
death rate or damageability of male’s systems in comparison
with corresponding female’s. In a course of Ontogeny the sex
ratio for many species of plants, animals and humans goes
down. It is related to the raised death rate and
damageability of male’s systems in comparison with female
ones at almost all Ontogeny stages and at all levels of
organization. Whether we study various species (the humans,
animals or plants), different levels of the organization (an
individual, organ, tissue or a cell) or stability to
different harmful factors of environment (low and high
temperature, starvation, poisons, parasites, diseases,
etc.)—anywhere the same picture is observed: the raised
death rate or damageability of male’s systems in comparison
with corresponding female’s.
Hamilton reviewed
death rate for 70 species, including such various forms of a
life, as nematodes, mollusks, crustaceans, insects, arachnoidea, birds, reptiles, fishes and mammals. According
to these data, for 62 species (89%) average life of males is
shorter, than females; for the majority of the remaining 11%
there is no difference, and only on occasion males live
longer, than females.
According to United Nations data, women
live longer than men in most countries. Higher death rate of
men is observed almost from all illnesses, with just a few
exceptions (pertussis, some gonococcal infections, etc.).
Males have also higher rates of unintentional as well as
intentional injuries as a consequence of more risky
behavior, aggression and job preference (police, fireman,
soldiers etc).
Summarizing, it is possible to tell, that the raised death
rate of males is a general biological phenomenon, it is
observed at plants, animals and humans for all levels of
organization from all extreme factors of environment.
The existing
theories
are not capable to explain the phenomenon of the raised
males’ death rate. The Evolutionary Theory of Sex considers raised death
rate and damageability of males as a favorable to a
population form of information contact with environment, a
payment for the new (ecological) information. This is
achieved due to the greater variation of male attributes.
In other words on all attributes, males should be more
various than females. Wider phenotypic variation of males
allows population “to pay” for the new information mostly by
male individuals.
Gamete Type and
Sex
With respect to sex, the conflict on
conservative and operative evolution tendencies was raised
at least twice. First time, when isogamy on the cell level
existed, the conflict requirements to their dimensions were
arisen. The operative task was to find another cell. In
order to accomplish this task, small size and mobility were
required. Conservative task was to preserve the formed
zygote, to supply nutrition and energy resources, protective
membranes etc., which is related to a big size. As a result
the gametes were differentiated by size and mobility on ova
and spermatozoids.
The differentiation by the gamete
type had the same problem. It was necessary to try recessive
genes before including them permanently in the genome. It
can be accomplished in autosomes in homozygous state, and in
gonosomes only in hemizigous state in heterogametic set of
chromosomes (XY). This is operative tendency. Heterozygous
gene combination in autosomes and in homogametic set (XX)
realize conservative tendency, because defective recessive
gene are not manifested.
In the process of evolution most
species had both operative subsystems (small gametes and
heterogametic constitution XY) in the male sex and both
conservative subsystems (large gametes and homogametic
constitution XX) in the female one. These are the species
with Drosophila type of gamety (Table).
Table Conservative and operative
subsystems on different
levels of organisation in mammals and birds.
Organization
level |
Heterogamety |
|
Male |
Female |
|
Organismic |
 |
 |
 |
 |
|
Cellular |
Spermatozoids |
Ova |
Spermatozoids |
Ova |
|
Chromosomal |
XY |
XX |
XX |
XY |
But, in the evolution of some species
the directions of these two differentiations did not match.
The conservative ova combined with the operative
heterogamety found themselves in the female sex, and
combination of operative spermatozoids with conservative
homogamety—in the male sex. These are species with
Abraxas type of gamety. So, from the presented point of
view, the Drosophila type of gamety is consistent,
but the Abraxas type of gamety is contradictory. This
can explain the fact that there are a lot more of
Drosophila species, assuming that the selection of type
was purely stochastic (independent of sex).
The role of gamety
type and polygamy
The effect of differential death rate
of sexes consists of two components (Figure
1). The first component, explained well by the theory
of imbalance of genes, is the contribution of gamety
type. The second component—is a contribution from
specialization of sexes at the population level. The
population effect arises in panmictic, freely crossed
population only, and is depends, probably, upon a degree of
polygamy. Therefore, to explain various death rates of sexes
one needs to take into account a gamety
type as well as mono- or polygamy of a given species.
 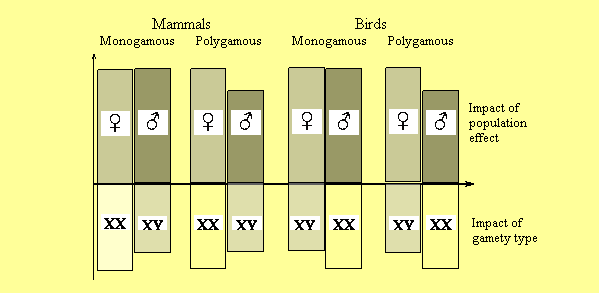
Figure 1
The impact of sex specialization at
the population level (the top part of figure) and the
contribution of gamety
type (the bottom part of figure) in differential death rate
of sexes. Population effect arises only in freely crossed
(polygamous) population, and is not valid for monogamous
species. For polygamous species the higher death rate
of males is common. A male sex and heterogametic
constitution (XY) are responsible for the operative task,
while
female sex and homogametic constitution (XX) – for the
conservative one.
The directions of these two differentiations
do not coincide in some species (for example, for birds).
Thus for polygamous species with female heterogamety,
the
population effect is directed against the effect of gamety,
while for polygamous species with male heterogamety
these
effects work in the same direction.
For a monogamous species the
population effects are minimal and the increased death rate
is observed for heterogametic sex (for males of Drosophila
type and females of Abraxas type). The facts are well
described by the theory of genes imbalance. For the
polygamous species of the Drosophila type, the population
effect is imposed upon the effect from gamety thus
strengthening it. This explains the maximal difference in
death rate observed for such species. For polygamous species
of Abraxas type, the population effect is directed against
the effect of gamety: heterogametic constitution leads to
the reduction of female longevity, while the population
effect—the male one. Therefore it is possible to expect,
that the difference in death rate for such species will be
less expressed, than for the polygamous species of the
Drosophila type with the same degree of polygamy. The
consequence from abovementioned statements is that
monogamous species of the Abraxas type should have higher
female mortality that actually was observed.
Therefore, the new approach explains
the observed pattern of differential mortality of sexes
well. Strictly speaking, for the realization of the
population mechanisms of sex specialization it is important
that male mortality should “precede” the female one. At an
adult age man’s death rate exceeds female. It partly can be
connected by that all “new” illnesses, illnesses of the
“century” or “civilization” (arteriosclerosis, hypertony,
cancer, AIDS, coronary diseases and schizophrenia), as a
rule, are illnesses of a male.
Existing theories—chromosomal
imbalance and metabolic—consider differential
mortality of sexes as a passive consequence of chromosomal
constitution or level of metabolism. They explain the
mechanism of a phenomenon and substitute the evolutionary
problem by genetic (imbalance theory) or physiological one
(metabolic theory). New theory considers increased mortality
of male sex as an active feature, increasing evolutionary
stability of the population. New approach clarifies
evolutionary meaning of differential mortality relating it
with different reaction norm of the sexes.
|