|
The Evolutionary Theory of
Sex:
High Male Mortality (II) The duration of life,
genotype and environment
 The duration of life, just as any
other character, is determined by the genotype and
environment. Let us imagine the following idealized
situations: (1) If the genotypic diversity in population is
eliminated (i.e. the clone of genetically identical
individuals) and unfavorable environmental factors are
removed (i.e. optimal conditions are created for a complete
realization of the single genotype) the life span of all the
individuals in such a clone will be similar, and the death
rate curve will be of a rectangular shape (variation
σ =0 Figure
2A).
(2) If instead of the clone a heterogeneous population is
taken in an optimum environment which permits realization of
all the genotypes, some variance in the life span will
appear which is conditioned only by genotypic variability
(variation σgen
Figure 2).
(3) Now if the clone again will be taken, but placed into a
real environment, the variance in the life span will be
conditioned by the environment (variation σenv
Figure 2).
(4) If a genetically heterogeneous population is placed
into real environment, the overall variation will include
both, the genotypic and environmental components (σ
= σgen + σenv)
Figure 2).
(5) Now let us imagine a situation when the death
rate is determined only stochastically, incidentally (by the
environment). Then the genotype will make no contribution to
the life span, and the latter will depend neither on age,
nor on health. In this case the death rate curve will have
an exponential pattern (Figure
2). The duration of life, just as any
other character, is determined by the genotype and
environment. Let us imagine the following idealized
situations: (1) If the genotypic diversity in population is
eliminated (i.e. the clone of genetically identical
individuals) and unfavorable environmental factors are
removed (i.e. optimal conditions are created for a complete
realization of the single genotype) the life span of all the
individuals in such a clone will be similar, and the death
rate curve will be of a rectangular shape (variation
σ =0 Figure
2A).
(2) If instead of the clone a heterogeneous population is
taken in an optimum environment which permits realization of
all the genotypes, some variance in the life span will
appear which is conditioned only by genotypic variability
(variation σgen
Figure 2).
(3) Now if the clone again will be taken, but placed into a
real environment, the variance in the life span will be
conditioned by the environment (variation σenv
Figure 2).
(4) If a genetically heterogeneous population is placed
into real environment, the overall variation will include
both, the genotypic and environmental components (σ
= σgen + σenv)
Figure 2).
(5) Now let us imagine a situation when the death
rate is determined only stochastically, incidentally (by the
environment). Then the genotype will make no contribution to
the life span, and the latter will depend neither on age,
nor on health. In this case the death rate curve will have
an exponential pattern (Figure
2).
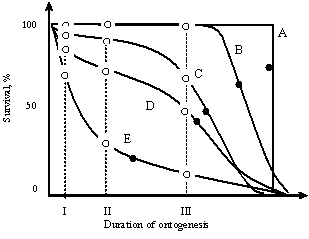
  Figure
2 Figure
2
Death rate curves determined to
varying degree by genotype and environment.
A—clone in ideal environment (σgen = 0, σenv = 0);
B—heterogeneous population in ideal environment (σgen
≠ 0, σenv = 0);
C— clone in real environment (σgen
= 0, σenv ≠ 0); D—heterogeneous population
in real environment (σgen ≠ 0, σenv ≠
0);
E—death rate is determined only stochastically.
I—average duration of intrauterine life;
II—average of appearance of reproductive capacity,
III—average age of loss of reproductive capacity,
● —average life span,
○ —average duration of Ontogeny stages.
It is evident that the curves (lB),
(1C), (1D) (as all the real cases met in life) are
intermediate between the extreme types (lA) (the death rate
is totally controlled by the genotype) and (lE) (it is under
total control of the environment), therefore they can be
presented as a superposition of these extreme types.
Consequently, as the population gets “emancipated” from the
environment, the pattern of its death rate approximates the
rectangular type and moves away from the exponential type,
and vice versa. Hence, the more optimum is the environment,
the nearer to the rectangular type is the death rate pattern
of the population; the more extreme it is—the closer to the
exponential type.
Now let us see how the transition of
the population from the optimum environment to the extreme
one affects the duration of Ontogeny and its stages. It
is evident that mortality strongly affects the average life
span and the post reproductive period and has practically no
effect on the average duration of periods of intrauterine
development, growth, and reproduction (Figure
2). This means
that a decrease in the average life span in extreme
environmental conditions is the result of the environmental
component of mortality and not the genotypic. Thus, the
application of the law of sexual dimorphism is a basis for
the hypothesis that genotypic average life span of men must
be greater than that of women. And if it was possible to
eliminate the effect of environment completely (to place the
population in an ideally optimal environment), then men
might possibly live longer than women. Such are the further
conclusions that might be drawn from the theory. Now let us
see what facts say about this.
Comparing the groups differently
“emancipated” from the environment, such as men and women of
one population, blacks and whites from one country, the
same country in the course of history, etc. (Comfort 1964),
one can see that the pattern of the death curve changes
regularly in the direction indicated by the theory. The more
is the group “emancipated” from the environment, the nearer
is its mortality curve to the rectangular type and vice
versa (see
Figure 3).
Greater “emancipation” of females as compared to males
results from a wider hereditary reaction norm of the women.
Greater “emancipation” of white population in the U.S.A.
compared to the African-American one, as well as of modern
population as compared with those of the former years is
conditioned by social and economical factors (nutrition,
medical service, etc.) (Figure
3).
 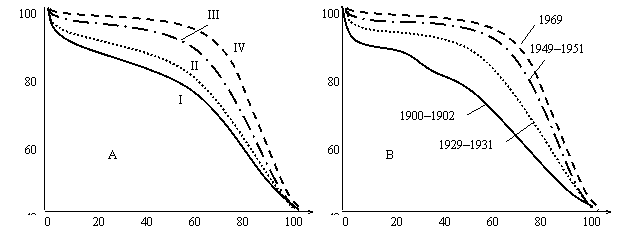
Figure 3
Death
rate curves (USA).
А: Number of survived per 100 000 (USA, 1939–1941):
I—African-American men;
II— African-American women; III—white men; IV— white women.
B: life span in USA in different years (Comfort, 1964).
Thus, the hypothesis suggested removes the contradictions
between the known correlations and the sexual dimorphism
rule. It explains why the average life span of women is
longer, while the
“champions” of longevity are men. For example, in the
central part of southern Transcaucasia, in the 1840s, 14 of
15 persons aged 110-140 were men. In Abkhazia in the 1920’s
all upper rungs of the age ladder were also occupied by men
(Arkhipov, 1978).
This hypothesis permits also to understand the seemingly
paradoxical fact that the phenomenon of long-living is
encountered in the population living under far from optimum
conditions (Comfort 1964, Davidovskij 1966).
If long-living is really conditioned by an increased
dispersion of the life span under extreme environmental
conditions, it should be closely related with stress as a
transmitter of ecological information in animals. It should
be accompanied by an increased death rate (the infants one
first of all) by an increased dispersion of other
characters, by the increased secondary sex ratio (birth of
boys) and by a rise of sexual dimorphism. The populations in
which long-living individuals are found should have the
death rate curve nearer to the exponential type,
characteristic of extreme conditions. While the death rate
curve of the population in an optimum environment should be
nearer to the rectangular type, where the long-living is not
observed.
Back to
Variation of Sexes and Reaction Norm ►
More about High Male Mortality:
Sexual Dimorphism and Evolution of Duration of
Ontogeny and its Stages. Geodakyan V. A. Doklady
Biological Sciences, 1982, v. 263, N 1-6, p. 174–177.
Translated from Doklady Akademii Nauk. SSSR, Genetika,
Vol. 263, No. 6, pp. 1475-1480, April, 1982.
|