Эволюционная
Теория
Пола:
Получение экологической информации от среды

Для регуляции популяционных параметров,
таких как плотность, темпы мутаций, соотношение полов,
дисперсия и половой диморфизм, нужна экологическая
информация. Особям популяции нужны какие-то датчики,
реагирующие на изменение экологических факторов среды.
Полученная информация потом преобразуется на организменном и
хромосомном уровнях.
Получение популяцией экологической
информации от среды заключается во-первых в том, что
изменение факторов среды может элиминировать самую
чувствительную к данному фактору часть особей популяции, в
результате естественного отбора. Bo-вторых,
изменение факторов среды, создав дискомфортные условия,
может полностью или частично отстранить от размножения
другую часть популяции, за счет полового отбора. В-третьих,
изменившаяся среда модифицирует выжившую часть популяции,
создавая морфо-физиологические, поведенческие и другие
ненаследуемые адаптации, за счет нормы реакции. Например в
холоде у животных укорачиваются хвосты, гуще становится мех,
утолщается подкожная жировая прослойка. Человек использует
пещеры, одежду, огонь.
Первые два процесса (элиминация и
дискриминация) вычеркивают какие-то генотипы из пула
воспроизводства. Третий процесс (модификация), наоборот,
позволяет каким-то генотипам сохраниться под прикрытием
модифицированного фенотипа и попасть в генофонд потомства.
То есть кого-то приходится ломать, убивать, отстранять, а
кого-то гнуть, “воспитывать”, переделывать. И то и другое
происходит, естественно, не в зоне комфорта, а в зонах
элиминации и дискомфорта.
Все эти процессы приводят, в конечном счете
к тому, что претерпевает изменения картина распределения
генотипов в двух смежных поколениях. Поколение n
получает от поколения п - 1 один спектр генотипов, а
передает поколению п + 1 другой. В этом
и заключается получение экологической информации от
среды. Стало быть, экологическая информация от среды в виде
мороза, жары, хищников, паразитов, воспринимается популяцией
в форме элиминации или модификации фенотипов. Это
меняет фенотипическое распределение популяции. Кроме
того, условия среды определяют репродуктивный успех,
подвергая дискриминации одни фенотипы и предоставляя
привилегии другим. В результате происходит изменение картины
генотипического распределения потомства.
Информативность особей на концах кривой
генотипического распределения (области E1
и E2)
максимальна, так как именно они могут вызвать максимальный
сдвиг генотипов в следующем поколении (Рис.
1). Чем дальше
генотип от моды, чем он оригинальнее, тем выше его
информативность. Однако самых оригинальных особей мало и их
вклад в генофонд потомства ничтожен.
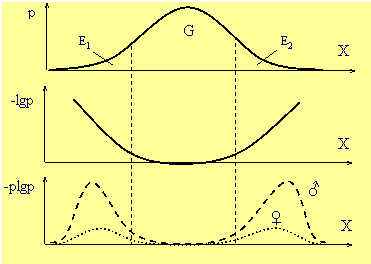
Рис.
1
Распределение фенотипов (p), их информативность (-lgp) и
информационная эффективность (-plgp) в популяции.
X—обобщенный признак (или фенотип), p—вероятность его
нахождения в популяции,
G—генетическая часть популяции,
E1, E2—экологические части.
Поэтому реальный вклад будет определяться
произведением степени оригинальности на концентрацию особей
с данным значением признака, а также на сечение их канала
связи с потомством.
Фенотипическая
дисперсия популяции
Чтобы популяция
“чувствовала” приближение фронта вредного фактора среды,
необходим контакт кривой смертности популяции с этим
фронтом. То есть за получаемую информацию популяции
приходится все время платить определенную жертву в виде
элиминации самых чувствительных к данному фактору особей.
При этом плата пропорциональна получаемой информации и тесно
связана с фенотипической дисперсией популяции. Если
фенотипическая дисперсия мала, то в стабильной среде нет
элиминации и нет информационного контакта популяции со
средой. В этом случае очередное внезапное изменение среды
может застать популяцию врасплох и погубить ее целиком.
Наоборот, если фенотипическая дисперсия очень велика, то
чрезмерно растет плата за новую информацию, а это
расточительно. Следовательно, существует некая оптимальная
для данной популяции в данной среде величина
фенотипической дисперсии, которая обеспечивает
заблаговременное получение необходимой информации при
минимальной плате за нее.
Если схематично
разделить популяцию на три части, как показано на
Рис.
2А,
то в привычных старых условиях среды, центральная часть
популяции пребывает в комфортных условиях по данному фактору
среды, а периферийные части—в условиях дискомфорта. При этом
периферийные части
E1
и E2
страдают от диаметрально противоположных (альтернативных)
факторов, например, если часть
E1
страдает от холода, то часть
E2—от
жары. Это означает, что даже в популяциях животных,
обитающих в тропиках, обезьян к примеру, какие-то экземпляры
погибают не только от жары, но и от холода, тогда как в
популяциях пингвинов или белых медведей, какие-то особи
погибают от тепла.
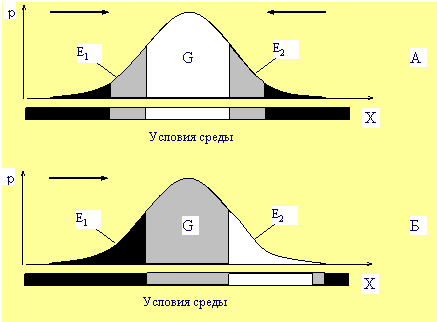
Рис. 2
Взаимоотношение популяции со средой в условиях
стабилизирующего (А) и движущего отбора (Б).
Зоны □—комфорта; ░—дискомфорта; █—элиминации.
→—направление давления отбора;
X—обобщенный признак (устойчивость к фактору среды),
p—вероятность (или концентрация) его
в популяции, G—генетическая часть популяции, E1,
E2—экологические части.
При изменении
условий среды, часть
E1
попадает в зону элиминации, часть
G—в
зону дискомфорта, а часть
E2—
в зону комфорта (Рис.
2Б).
Особи класса E2,
которые в обычных условиях страдали от дискомфорта, попав
наконец, в комфортные условия, бурно размножаются и “уводят”
популяцию от наступающего вредного фактора среды.
То есть,
информационная ценность особей разных классов разная.
Центральный класс, максимально приспособленный к старым
условиям и поэтому ориентированный как бы на прошлое,
“работает” больше на сохранение имеющейся генетической
информации, реализует больше наследственную тенденцию—сохранения.
Периферийные классы
E1
и E2
оба плохо приспособленные к старым условиям среды, новые
условия встречают по разному: особи класса
E1
гибнут, а класса
E2
становятся самыми приспособленными. Поэтому можно сказать,
что классы E1
и E2
“работают” больше на изменение, реализуют больше
экологическую тенденцию, связь со средой и ориентированы
больше на будущее. Иными словами, в популяции существует
специализация, своего рода “разделение труда” по связи со
средой (изменчивость) и по связи с предшествующими
поколениями (наследственность).
Механизмы регуляции параметров популяции
(стресс, гормоны, количество пыльцы)
Два пола—два
потока информации
|