|
Половые хромосомы: для чего
они? (I)

Новая концепция
использует идею асинхронной эволюции для объяснения
существования и функциональной роли половых
хромосом (ПХ).
В 1965 г. В. Геодакян
теоретически предсказал, что: “в хромосомном наборе половые
хромосомы выполняют роль оперативной памяти, а аутосомы
(А)—постоянной, поэтому половые хромосомы (в первую
очередь Y-хромосома) являются “воротами” для изменчивости в
наследственность” (Геодакян,1965). Y-хромосома делает это в
основном при помощи мутаций индуцированных и направляемых
экологической информацией чем спонтанных мутаций.
Новые гены сначала
появляются в Y-хромосоме и потом
через много поколений переходят через Х-хромосому в
аутосомы.
Каждый новый ген,
прежде чем попасть в аутосомы, проходит двойную проверку в
половых хромосомах: сначала в Y-хромосоме,
и потом в Х-хромосоме. Но поскольку рецессивные Х-гены
проявляются только у мужского пола, то отбору
подвергается только он. В этом состоит эволюционное значение
половой дифференциации и основное преимущество полового
диморфизма.
Эволюционная логика аутосом и половых
хромосом.
Аутосомы
являются консервативной памятью генома (аналог женского пола
в популяции) и нацелены на его сохранение. Они хранят
стабильные гены, общие для обоих полов. Эволюционно это
самые старые хромосомы, содержащие фундаментальную видовую
информацию. Они выполняют наиболее древние программы
репродукции и рекомбинации. Передаваясь
случайно, они перемешиваются в каждом поколении,
обеспечивая максимальное разнообразие генотипов, то
есть
наилучшим образом реализуют программы полового процесса, по
которым наивысшие достижения у гермафродитов. В этом смысле
они скорее “рекомбинационные”.
Половые хромосомы
являются оперативной памятью или экспериментальной
подсистемой генома и нацелены на его изменение
(аналог мужского пола в популяции). Так же как новый признак
не появляется в женском фенотипе, не пройдя проверку в
мужском, так и новый ген не появляется в аутосомах не пройдя
проверку в половых хромосомах.
Основная роль половых
хромосом не репродуктивная—определение пола, как
считали раньше, а эволюционная—создание
двух сдвинутых по времени форм (женской и мужской)
для экономной эволюции (даже в ущерб размножению!),
не обеспечение соотношения полов = 1:1
и его постоянства, а наоборот, его изменение
(регуляция).
Половые хромосомы
значительно “моложе” аутосом. Запуская и выполняя программу
дифференциации, они формируют в популяции
консервативно-оперативные подсистемы. Для этого они снабжают
более моногамный пол широкой нормой реакции, а более
полигамный — узкой независимо от типа гаметности. В них
находятся, главным образом, эволюционирующие гены, как
приобретаемые, так и утрачиваемые. Их деятельность, будучи
направлена против программы рекомбинации
(запрет комбинаций ♂♂ и ♀♀), ухудшает вдвое разнообразие
генотипов, возможное в результате полового процесса и в этом
смысле они скорее “антиполовые”, чем “половые”. Поэтому
логичнее было бы, с учетом их роли, назвать их —
“эволюционными”.
Алгоритмы хромосом и их сопоставление с фазами эволюции
В информационном поведении хромосом можно выделить
“вертикальные” и “горизонтальные” алгоритмы. “Вертикальные”
— это передача самих хромосом из поколения в поколение.
“Горизонтальные” — поступление информации от среды, передача
её между хромосомами и элиминация, в виде генетических
процессов мутагенеза, кроссинговера, транслокаций, переноса
эписомами, вирусами, плазмидами, мобильными генами и т.д.
Поведение хромосом определяют прежде всего три вертикальных
алгоритма. 1. Стохастический, когда хромосомы
гомологичной пары попадают сыну или дочери чисто случайно.
Этим алгоритмом передаются аутосомы и (считается)
Х-хромосомы гомогаметного пола. 2. Ипси-алгоритм,
когда хромосома переходит от родителя только потомку того
же пола. Это алгоритм
Y-хромосомы.
3. Контра-алгоритм осуществляет переход от
родителя к потомку противоположного пола. Это
алгоритм Х-хромосомы гетерогаметного пола.
Стохастический
алгоритм имеет дело только с общей для обоих полов
генетической информацией. Перемешивая гены при каждом
оплодотворении, он максимизирует генетическое разнообразие,
поэтому на его основе не может быть генотипический
половой диморфизм. Это самый
древний алгоритм, который существовал еще до появления
дифференциации.
Нестохастические
алгоритмы возникли с появлением половых хромосом и имеют
дело с информацией, разной у мужского и женского пола.
Ипси-алгоритм, действуя в пределах одного пола,
может создавать генотипический
половой диморфизм и менять его величину.
Он — инициатор программы дифференциации. Контра-алгоритм,
также как и стохастический, переносит информацию от одного
пола другому, поэтому он тоже уменьшает разницу между
полами. В отличие от стохастического алгоритма, он не сводит
половой диморфизм к нулю,
а сохраняет его постоянство.
Сочетание ипси-контра алгоритмов позволяет создавать и
сохранять определенную разницу (“дистанцию”) между
подсистемами и менять её в зависимости от условий среды.
При этом контра-алгоритм выступает как стабилизатор
(отрицательная обратная связь), а ипси-алгоритм — как
регулятор (положительная обратная связь).
Y-хромосома —
связующее звено между ядром и средой (цитоплазмой,
митохондриями), “ворота” генома для новой информаци. Она
трансформирует экологическую информацию в генетическую, т.е.
создает новые гены (мутации), поэтому ее можно назвать
“экологической”. Она содержит “завтрашние” гены, является
инициатором, акселератором и регулятором асинхронии,
полового дихрономорфизма (эволюционной и морфологической
“дистанции” между полами, в том числе и по норме реакции);
представляет собой первый “испытательный полигон” и
“карантинный изолятор” для новых генов.
Контра-Х-хромосома (cX) —
является переносчиком генов, связующим звеном между
Y-хромосомой
и женским геномом (“транспортная” хромосома). В филогенезе
функционирует как стабилизатор, релаксатор и ликвидатор
полового дихрономорфизма. Она также представляет собой
второй полигон, где молодые гены, находящиеся в гемизиготном
состоянии у мужского пола и в гетерозиготном—у женского,
проходят испытания в онтогенезе.
Ипси-Х-хромосома (iX)
— должна быть связана прежде всего с определением нормы
реакции женского пола в зависимости от типа полигамии. В ней
должна быть больше доля модификационных генов,
количественных признаков. Она переносит новые гены в
аутосомы и удаляет из них старые гены. Определенный ее
участок содержит сугубо женские “вчерашние” гены, то есть
несет атавистическую информацию. Таким образом половые
хромосомы осуществляют как хранение, так и перенос
информации.
Эволюционный маршрут генов по хромосомам
Филогенетический путь переноса информации
следующий:
|
Среда |
|
_______________
Хромосомы ________________ |
|
↓ |
|
│ |
|
|
|
|
|
|
|
|
|
|
|
│ |
| цитоплазма |
→ |
Y |
→ |
cXm |
→ |
cXf |
→ |
A |
→ |
iXf |
→ |
cXf |
→ |
cXm |
| |
Появление новых генов
—> |
│ |
—>
Смерть ненужных генов |
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
m – самцы, f – самки |
При частичной конъюгации Х и
Y-хромосом
как у животных, так и у растений, только часть генов
находится в конъюгирующем участке (Рис.). Следовательно,
“вход” и “выход”
Y-хромосомы
должны быть удалены друг от друга. Должно пройти какое-то
время, чтобы ген перемещаясь вдоль Y-хромосомы попал в
участок конъюгирующий с Х-хромосомой. Затем неравным
кроссинговером между Y и Х-хромосомами он передается в
женский геном.
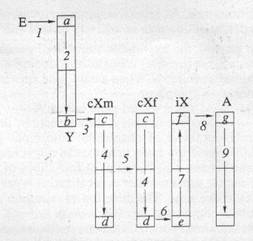
Рис.
Гипотетическая схема маршрута нового гена по половым
хромосомам в дивергентной фазе эволюции признака.
E
—среда
(цитоплазма). Участки хромосом:
a,
c,
e,
g
- входы хромосом, не участвующие в кроссинговере;
b,
d,
f
- выходы хромосом, участвующие в неравном кроссинговере и
транслокации генов на аутосомы. Переходы генов: 1 - из
cреды
в
Y-хромосому
(мутагенез); 2 - по длине
Y-хромосомы
из неконъюгирующего (а) участка в конъюгирующий (б); 3 -
неравный кроссинговер
Y®
cХм;
4 - по длине
cХ-хромосомы
в мужском и женском геноме; 5 - вертикальный
cХ-алгоритм
(отец
®
дочь); 6 - неравный кроссинговер:
cХм®iХ;
7 - по длине
iХ-хромосомы;
8 - транслокация (плазмиды, вирусы)
iХ-хромосома
®
аутосомы . 9 - по длине аутосом.
В Х-хромосоме, видимо, так же, как и в
Y,
“вход” и “выход” удалены друг от друга, поэтому,
передвигаясь вдоль Х-хромосомы, “молодой” ген проходит в
мужском геноме вторые испытания, будучи в гемизиготном
состоянии (Рис.). Значит каждый новый ген, прежде чем
попасть в аутосомы, проходит двойную проверку в половых
хромосомах: сначала в
Y-хромосоме,
потом в Х-хромосоме. Но поскольку рецессивные Х-гены
проявляются по сути только у мужского пола, то отбору
подвергается только он.
Половые хромосомы: для чего
они? (II)
►
|